Apakah perbezaan antara alternatif klasik dan laluan lektin
The Perbezaan utama antara alternatif klasik dan laluan lektin adalah bahawa permulaan laluan klasik berlaku melalui pengikatan kompleks antigen-antibodi ke protein C1q, dan permulaan laluan alternatif berlaku melalui pengikatan C3b ke permukaan asing, sementara permulaan jalur lektin berlaku melalui mannose-mengikat lectin.
Laluan pelengkap atau pelengkap litar adalah sebahagian daripada sistem imun yang meningkatkan keupayaan sel fagositik dan antibodi untuk memusnahkan dan membersihkan mikroba dan sel yang rosak dari organisma, menggalakkan keradangan, dan menyerang membran sel patogen. Laluan pelengkap dihasilkan oleh sistem imun yang semula jadi dan adaptif. Sistem ini terdiri daripada protein kecil yang disintesis oleh hati dan beredar dalam darah dalam bentuk yang tidak aktif. Protein atau prekursor ini diaktifkan dalam laluan pelengkap. Terdapat tiga jenis laluan pelengkap: laluan klasik, laluan alternatif, dan laluan lektin.
Kandungan
1. Gambaran Keseluruhan dan Perbezaan Utama
2. Apakah laluan klasik
3. Apakah laluan alternatif
4. Apakah laluan lektin
5. Persamaan - Laluan Alternatif dan Lektin Klasik
6. Klasik vs alternatif vs jalur lektin dalam bentuk jadual
7. Ringkasan - Jalur lektin alternatif vs klasik vs
Apakah laluan klasik?
Laluan Klasik adalah salah satu daripada tiga laluan yang mengaktifkan sistem pelengkap. Sistem pelengkap adalah sebahagian daripada sistem imun. Kompleks antigen-antibodi bersama dengan isotypes antibodi IgG dan IgM memulakan sistem pelengkap. Sel apoptotik, sel nekrotik, dan protein fasa akut juga mengaktifkan laluan klasik.
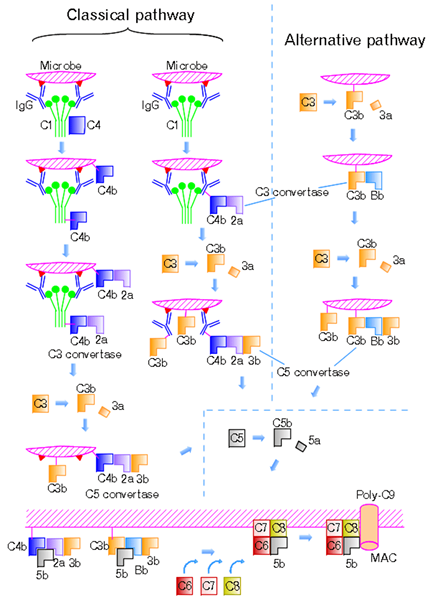
Rajah 01: Laluan Klasik dan Laluan Alternatif
Laluan ini dimulakan oleh pengikatan kompleks antigen-antibodi kepada protein C1Q; Kawasan globular C1Q mengiktiraf dan mengikat ke rantau FC antibodi IgG dan IgM isotypes. Mereka juga mengikat protein permukaan bakteria dan virus, sel apoptosis, dan protein fasa akut. Semasa ketiadaan faktor pengaktifan, C1Q menjadi sebahagian daripada kompleks C1 yang tidak aktif, yang terdiri daripada enam molekul C1Q, dua molekul C1R, dan dua molekul C1S. Pengikatan C1Q membawa kepada perubahan konformasi dan pengaktifan protease serine C1R. Ini mengaktifkan dan mengekang c1 protease serine. C1s kemudian memecah C4 ke C4A dan C4B dan C2 ke C2A dan C2B. C4B membantu dalam pembentukan C3 convertase, C4BC2A. Convertase C3 mempunyai keupayaan untuk memecahkan C3 ke C3a dan C3b, yang merupakan faktor penting untuk reaksi enzimatik seterusnya. C3V mengikat kepada penukaran C3 untuk membentuk penukaran C5, C4B2A3B, manakala C3A merekrut sel keradangan melalui. Ini dikenali sebagai anafilatoksin. C5 Convertase Cleaves C5 menjadi C5 A dan C5b. C5B menggabungkan dengan komponen terminal lain untuk membentuk Kompleks Serangan Membran (MAC). Ini membawa kepada lisis bakteria invasif dengan memasukkan ke dalam membran sel sasaran, mewujudkan liang berfungsi.
Apakah laluan alternatif?
Laluan alternatif adalah salah satu daripada tiga laluan yang opsonize dan memusnahkan patogen. Virus, kulat, bakteria, parasit, immunoglobulin A dan polysaccharides mengaktifkan laluan alternatif dan membentuk mekanisme pertahanan penting yang bebas daripada sistem imun. Protein C3b mencetuskan laluan ini, dan protein ini secara langsung mengikat mikroba. Bahan asing dan tisu yang rosak juga mencetuskan laluan alternatif. Oleh kerana C3b adalah percuma dan berlimpah dalam plasma, ia mempunyai keupayaan untuk mengikat sel tuan rumah atau permukaan patogen. Protein pengawalseliaan yang berbeza mengambil bahagian dalam pencegahan pengaktifan pelengkap pada sel tuan rumah.
Pelengkap Reseptor 1 (CR1) dan faktor pereputan (DAF) bersaing dengan Faktor B untuk mengikat dengan C3b di permukaan sel dan keluarkan BB dari kompleks C3BBB. C3b ke dalam bentuk yang tidak aktif, IC3b, oleh protease plasma yang dipanggil faktor pelengkap 1 menghalang pembentukan penukaran C3. Faktor pelengkap 1 memerlukan cofactor protein mengikat C3B seperti faktor H, Cr1, atau membran cofactor proteolisis. Faktor H menghalang pembentukan penukaran C3 dengan bersaing dengan faktor B untuk mengikat dengan C3b. Ini juga mempercepatkan kerosakan penukaran C3. CFHR5, yang merupakan faktor pelengkap H protein yang berkaitan 5, mempunyai keupayaan untuk mengikat bertindak sebagai cofactor untuk faktor 1 dan mempercepatkan aktiviti pembusukan, dan mengikat C3b pada sel tuan rumah.
Apakah laluan lektin?
Laluan lektin adalah jenis reaksi lata dalam sistem pelengkap. Selepas pengaktifan laluan ini, tindakan C4 dan C2 menghasilkan protein pelengkap yang diaktifkan ke bawah lata. Laluan ini tidak mengiktiraf antibodi yang terikat pada sasarannya dan memulakan dengan lektin mannose (MBL) atau ficolin yang mengikat gula-gula tertentu. MBL ini mengikat gula seperti mannose dan glukosa dengan kumpulan OH dalam kedudukan terminal pada karbohidrat atau komponen glikoprotein bakteria, kulat, dan beberapa virus.
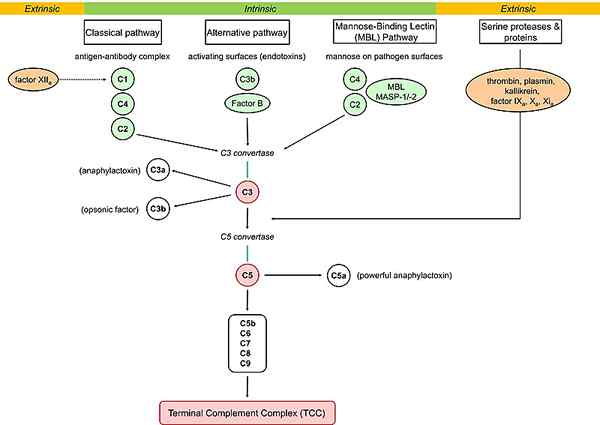
Rajah 02: pelengkap laluan
MBL, juga dikenali sebagai protein mengikat mannose, mampu memulakan sistem pelengkap dengan mengikat permukaan patogen. Multimer MBL membentuk kompleks dengan protease serine (mannose mengikat lektin serine yang berkaitan dengan protease: MASP1, MASP2 dan MASP3) yang merupakan zymogens protein. Mereka serupa dengan C1R dan C1 di laluan lain. MASP1 dan MASP2 mengaktifkan komponen Cleave C4 dan C2 ke C4A, C4B, C2A, dan C2B. C4B cenderung mengikat membran sel bakteria. Sekiranya ia tidak diaktifkan, ia menggabungkan dengan C2A untuk membentuk penukaran C3 klasik yang menentang penukaran C3 alternatif. C4A dan C2B bertindak sebagai sitokin yang kuat. C4A menyebabkan degranulasi sel mast dan basofil, dan C2B meningkatkan kebolehtelapan vaskular.
Apakah persamaan antara laluan alternatif dan lektin klasik?
- Laluan klasik, alternatif, dan lektin diaktifkan oleh litar tindak balas yang membawa kepada kompleks serangan membran.
- Mereka adalah sebahagian daripada sistem imun.
- Setiap laluan mempunyai protein yang unik untuk permulaan.
- Mereka diaktifkan oleh isotip tertentu antibodi yang terikat kepada antigen
Apakah perbezaan antara alternatif klasik dan laluan lektin?
Permulaan laluan klasik berlaku melalui pengikatan kompleks antigen-antibodi kepada protein C1Q. Inisiasi laluan alternatif berlaku melalui pengikatan C3b ke permukaan asing, sementara permulaan jalur lektin berlaku melalui lektin yang mengikat mannose. Oleh itu, ini adalah perbezaan utama antara laluan alternatif dan lektin klasik. Peranan jalur klasik adalah bahawa ia bertindak sebagai lengan effector imuniti penyesuaian manakala laluan alternatif dan lektin berfungsi dalam imuniti semula jadi. Selain itu, pengaktifan C4 dan C2 dalam laluan klasik adalah C1, dan dalam laluan lektin adalah MASP-2, sementara tidak ada pengaktifan C4 dan C2 dalam laluan alternatif.
Infographic di bawah membentangkan perbezaan antara alternatif klasik dan laluan lektin dalam bentuk tabular untuk perbandingan sampingan.
Ringkasan -Jalur lektin alternatif vs klasik vs
Permulaan laluan klasik berlaku melalui pengikatan kompleks antigen-antibodi kepada protein C1Q. Inisiasi laluan alternatif berlaku melalui pengikatan C3B ke permukaan asing sementara permulaan laluan lektin berlaku melalui lektin yang mengikat mannose. Jadi, ini adalah perbezaan utama antara alternatif klasik dan laluan lektin.
Rujukan:
1. "Laluan Klasik."Biolab kreatif.
2. Fujita, Teizo. "Evolusi laluan pelengkap lektin dan peranannya dalam imuniti semula jadi."Nature News, Nature Publishing Group.
Ihsan gambar:
1. "Melengkapkan laluan."Oleh tossh_eng - https: // muat naik.Wikimedia.org/wikipedia/commons/b/b0/jalan pelengkap.PNG (CC BY-SA 4.0) melalui Commons Wikimedia
2. "Rajah 1.4. Pelengkap pengaktifan "oleh Hofman.Martijn - Kerja Sendiri (CC BY -SA 4.0) melalui Commons Wikimedia