Perbezaan antara pembaikan tidak sepadan dan pembaikan pengasingan nukleotida
Perbezaan Utama - Pembaikan Mismatch vs Pembaikan Pengecualian Nukleotida
Puluhan dan ribuan kerosakan DNA berlaku di sel sehari. Ia mendorong perubahan kepada proses sel seperti replikasi, transkripsi serta daya maju sel. Dalam sesetengah kes, mutasi yang disebabkan oleh kerosakan DNA ini boleh menyebabkan penyakit yang merosakkan seperti kanser dan sindrom yang berkaitan dengan penuaan (Ex: Progeria). Tidak kira kerosakan ini, sel memulakan mekanisme pembaikan lata yang sangat teratur yang disebut tindak balas kerosakan DNA. Beberapa sistem pembaikan DNA telah dikenalpasti dalam sistem selular; Ini dikenali sebagai Pembaikan Pengasingan Asas (BER), Pembaikan Tidak Taksiran (MMR), Pembaikan Pengasingan Nukleotida (NER), Pembaikan Break Strand Double. Pembaikan Pengecualian Nukleotida adalah sistem yang sangat serba boleh yang mengiktiraf lesi DNA distorsi helix yang besar dan menghilangkannya. Sebaliknya, pembaikan tidak sepadan menggantikan pangkalan yang tidak diperbadankan semasa replikasi. Perbezaan utama antara pembaikan tidak sepadan dan pembaikan pengasingan nukleotida ialah Pembaikan Pengecualian Nukleotida (NER) digunakan untuk menghilangkan dimer pyrimidine yang dibentuk oleh penyinaran UV dan lesi helix yang besar yang disebabkan oleh penambahan kimia manakala sistem pembaikan tidak sepadan memainkan peranan penting dalam membetulkan asas yang tidak disengajakan yang telah melarikan diri dari enzim replikasi (DNA molymerase 1). Sebagai tambahan kepada pangkalan yang tidak sesuai, protein sistem MMR juga boleh membaiki gelung sisipan/ penghapusan (IDL) yang merupakan hasil slip polimerase semasa replikasi urutan DNA berulang.
Kandungan
1. Gambaran Keseluruhan dan Perbezaan Utama
2. Apakah pembaikan tidak sepadan
3. Apakah pembaikan pengasingan nukleotida
4. Perbandingan sampingan - Pembaikan tidak sepadan dengan pembaikan pengasingan nukleotida
5. Ringkasan
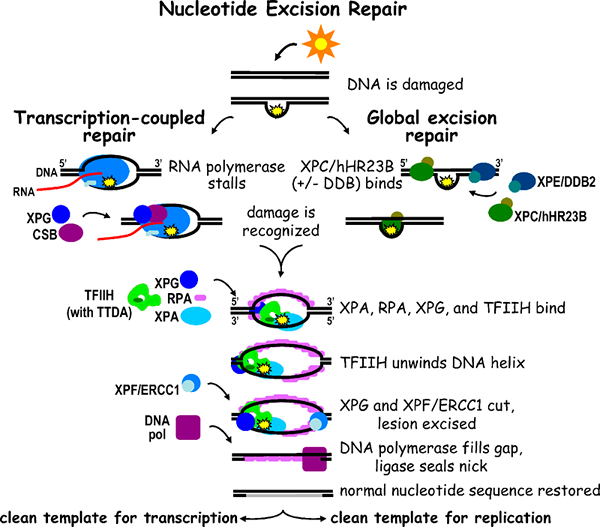
Apakah pembaikan pengasingan nukleotida?
Ciri pembaikan pengasingan nukleotida yang paling terkenal ialah pembaikan kerosakan nukleotida yang diubahsuai yang disebabkan oleh gangguan yang ketara dalam helix double DNA. Ia diperhatikan dalam hampir semua organisma yang telah diperiksa sehingga kini. UVR A, UVR B, UVR C (Excinucleases) UVR D (Helicase) adalah enzim yang paling terkenal yang terlibat dalam NER yang mencetuskan pembaikan DNA dalam model organisme Ecoli. Kompleks enzim multi-subunit UVR ABC menghasilkan polipeptida UVR A, UVR B, UVR C. Gen yang dikodkan untuk polipeptida yang disebutkan di atas adalah UVR A, UVR B, UVR C. Enzim UVR A dan B secara kolektif mengiktiraf kerosakan yang disebabkan oleh distorsi yang disebabkan oleh helix double DNA seperti dimmers pyrimidine kerana penyinaran UV. UVR A adalah enzim ATPase dan ini adalah tindak balas autokatalik. Kemudian UVR A meninggalkan DNA manakala kompleks UVR BC (nukleus aktif) memecah DNA di kedua -dua belah kerosakan yang dikatalisis oleh ATP. Satu lagi protein yang dipanggil UVR D yang dikodkan oleh gen UVRD adalah enzim helikase II yang melepaskan DNA yang disebabkan oleh pelepasan segmen DNA yang rosak terkandas tunggal. Ini meninggalkan jurang dalam helix DNA. Selepas segmen yang rosak telah dikeluarkan, jurang nukleotida 12-13 kekal di helai DNA. Ini dipenuhi oleh enzim polimerase DNA I dan nick dimeteraikan oleh ligase DNA. ATP diperlukan pada tiga langkah reaksi ini. Mekanisme NER dapat dikenal pasti dalam manusia seperti mamalia. Pada manusia, keadaan kulit yang disebut xeroderma pigmentosum disebabkan oleh dimer DNA yang disebabkan oleh penyinaran UV. Gen XPA, XPB, XPC, XPD, XPE, XPF, dan XPG menghasilkan protein untuk menggantikan kerosakan DNA. Protein gen XPA, XPC, XPE, XPF, dan XPG mempunyai aktiviti nukleus. Sebaliknya, protein gen XPB dan XPD menunjukkan aktiviti helikase yang analog ke UVR D dalam E coli.
Rajah 01: Pembaikan pengasingan nukleotida
Apakah pembaikan tidak sepadan?
Sistem Pembaikan Mismatch dimulakan semasa sintesis DNA. Walaupun dengan subunit € berfungsi, polimerase DNA III membolehkan penggabungan nukleotida yang salah untuk sintesis setiap 108 Pasangan asas. Protein pembaikan tidak sepadan mengiktiraf nukleotida ini, mengeluarkannya dan menggantinya dengan nukleotida yang betul yang bertanggungjawab untuk tahap ketepatan akhir. Metilasi DNA penting bagi protein MMR untuk mengenali helai induk dari helai yang baru disintesis. Metilasi adenine (a) nukleotida dalam motif GATC dari helai yang baru disintesis sedikit ditangguhkan. Sebaliknya, nukleotida adenine strand induk dalam motif GATC telah metilated. Protein MMR mengiktiraf helai yang baru disintesis dengan perbezaan ini dari helai induk dan memulakan pembaikan tidak sepadan dalam helai yang baru disintesis sebelum ia mendapat metilasi. Protein MMR mengarahkan aktiviti pembaikan mereka untuk mengeluarkan nukleotida yang salah sebelum helai DNA yang baru direplikasi mendapat metilasi. Enzim mut h, mut l dan mut s yang dikodkan oleh gen mut h, mut l, mut s memangkinkan reaksi ini dalam ecoli. Protein Mut S mengiktiraf tujuh daripada lapan pasangan asas yang mungkin tidak sepadan kecuali C: C, dan mengikat di tapak ketidakcocokan dalam DNA dupleks. Dengan atps terikat, mut l dan mut s menyertai kompleks kemudian. Kompleks translocates beberapa ribu pasangan asas sehingga ia mendapati motif GATC hemimethylated. Aktiviti nukleus tidak aktif protein Mut H diaktifkan apabila ia mendapati motif GATC hemimetilasi. Ia memecahkan helai DNA yang tidak metilat yang meninggalkan 5 'Nick pada G nukleotida motif GATC yang tidak dimetilkan (helai DNA yang baru disintesis). Kemudian helai yang sama di sisi lain ketidakcocokan disusun oleh mut h. Sepanjang langkah-langkah, tindakan kolektif UVR d protein helikase, mut u, ssb dan exonuclease i mengeluarkan nukleotida yang salah dalam DNA tunggal terkandas. Jurang yang terbentuk dalam pengasingan diisi oleh polimerase DNA III dan dimeteraikan oleh ligase. Sistem yang serupa dapat dikenal pasti pada tikus dan manusia. Mutasi manusia HMLH1, HMSH1, dan HMSH2 terlibat dalam kanser kolon nonpolyposis keturunan yang menghancurkan sel sel kolon.
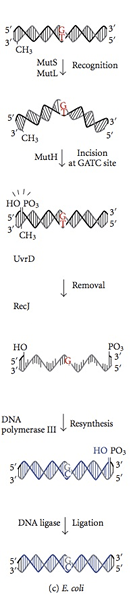
Rajah 02: Pembaikan tidak sepadan
Apakah perbezaan antara pembaikan tidak sepadan dan pembaikan pengasingan nukleotida?
Pembaikan Mismatch vs Pembaikan Pengasingan Nukleotida | |
| Sistem pembaikan tidak sepadan berlaku semasa replikasi selepas. | Ini terlibat dalam mengeluarkan dimer pyrimidine kerana anda.V Irradiasi & luka DNA lain disebabkan oleh penambahan kimia. |
| Enzim | |
| Ia dipangkin oleh mut s, mut l, mut h, uvr d, ssb dan exonuclease i. | Ia dipangkin oleh enzim UVR A, UVR B, UVR C, UVRD. |
| Metilasi | |
| Sangat penting untuk memulakan reaksi. | Metilasi DNA tidak diperlukan untuk memulakan reaksi. |
| Tindakan enzim | |
| Mut h adalah endonuclease. | Uvr b dan uvr c adalah exonucleases. |
| Kesempatan | |
| Ini berlaku secara khusus semasa replikasi. | Ini berlaku apabila terdedah kepada anda.V atau mutagen kimia, bukan semasa replikasi |
| Pemuliharaan | |
| Ia sangat konservasi | Ia tidak sangat konservasi. |
| Pengisian jurang | |
| Ia dilakukan oleh polimerase DNA III. | Ia dilakukan oleh polimerase DNA i. |
Ringkasan - Pembaikan Mismatch vs Pembaikan Pengasingan Nukleotida
Pembaikan Mismatch (MMR) dan Pembaikan Pengasingan Nukleotida (NER) adalah dua mekanisme yang berlaku di dalam sel untuk membetulkan kerosakan DNA dan gangguan yang disebabkan oleh pelbagai ejen. Ini secara kolektif dinamakan sebagai mekanisme pembaikan DNA. Pembaikan Pembaikan Pengasingan Nukleotida Pembaikan kerosakan nukleotida yang diubahsuai, biasanya kerosakan besar DNA double helix yang berlaku disebabkan oleh pendedahan kepada u.V penyinaran dan penambahan kimia. Protein pembaikan tidak sepadan mengiktiraf nukleotida yang salah, mengeluarkannya dan menggantikannya dengan nukleotida yang betul. Proses ini bertanggungjawab untuk tahap ketepatan akhir semasa replikasi.
Rujukan:
1.Cooper, Geoffrey m. "Pembaikan DNA."Sel: Pendekatan Molekul. Edisi ke -2.U.S. Perpustakaan Perubatan Negara, 01 Jan. 1970. Web. 09 Mac. 2017.
2."Mekanisme dan fungsi pembaikan tidak sepadan DNA."Penyelidikan Sel. U.S. Perpustakaan Perubatan Negara, n.d. Web. 09 Mac. 2017.
Ihsan gambar:
1. "Pembaikan Pengasingan Nukleotida-Journal.pbio.0040203.G001 "oleh Jill O. Fuss, Priscilla k. Cooper - (CC oleh 2.5) Melalui Wikimedia Commons
2. "Ecoli Pembaikan Mismatch DNA" oleh Kenji Fukui - (CC oleh 4.0) melalui Commons Wikimedia